Per una sintesi si rimanda all’ultimo paragrafo.
Introduzione
Nel mio precedente articolo sullo studio pubblicato dal gruppo di Fluge e Mella, mi sono limitato a riportare i loro risultati sulla analisi dei 20 amminoacidi standard nel sangue dei pazienti ME/CFS. Riassumendo:
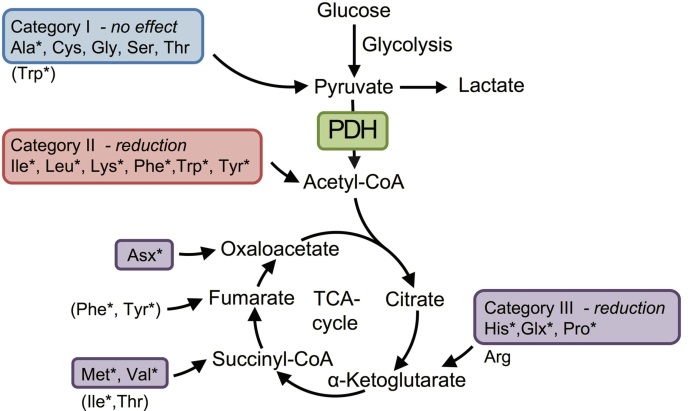
- nelle donne con ME/CFS gli amminoacidi che si trasformano in acetil-CoA e quelli che alimentano direttamente il ciclo dell’acido citrico (ciclo TCA) sono ridotti;
- nei maschi non si ha questa riduzione degli amminoacidi nel sangue, ma si ha un aumento della 3-metilistidina (3-MHis), che suggerisce che i muscoli vengono smembrati (catabolizzati) per alimentare il ciclo del TCA. Nelle donne questo fenomeno è assente.
Gli autori hanno formulato allora la seguente ipotesi:
- i pazienti ME/CFS (sia maschi che femmine) utilizzano gli amminoacidi per alimentare i mitocondri (il ciclo del TCA avviene dentro i mitocondri) più di quanto facciano i controlli sani.
I mitocondri – in condizioni normali – usano come fonte principale di alimentazione lo zucchero. Lo zucchero – trasformato in piruvato fuori dai mitocondri (nella glicolisi) – viene ulteriormente trasformato in acetil-CoA dal PDH, e quindi alimenta il ciclo del TCA (vedi figura 1).
Ciò posto, una possibile spiegazione per l’aumentato consumo degli amminoacidi da parte dei pazienti ME/CFS è che:
- l’attività dell’enzima piruvato deidrogenasi (PDH) sia ridotta – per qualche motivo – e quindi l’approvigionamento di acetil-CoA è deficitario. In questo contesto, i mitocondri bruciano gli amminoacidi per produre energia, al posto dell zucchero.
Espressione genica
Per verificare l’ipotesi di un ridotto funzionamento del piruvato deidrogenasi (PDH), gli autori sono passati a un secondo gruppo di analisi, che sono l’oggetto del presente articolo. In particolare, il gruppo norvegese ha misurato l’RNA messaggero (l’espressione genica) di diversi geni che sono coinvolti nella regolazione della attività del PDK. L’espressione genica è stata misurata in un gruppo di cellule facilmente accessibili, ovvero le cellule mononucleari del sangue periferico (PBMC). In pratica le PBMC sono: linfociti (cellule B, T, NK) e monociti (macrofagi). Ebbene, diversi geni coinvolti nella inibizione del piruvato deidrogenasi risultano sovra espressi, tanto nei maschi che nelle femmine con ME/CFS. Vediamo quali.

Piruvato deidrogenasi chinasi
L’enzima piruvato deidrogenasi chinasi (PDK) ha come principale funzione quella di inibire il PDH. Dunque una buona domanda da farsi è: quanto PDK è espresso nelle cellule delle persone con ME/CFS? Se l’attività del PDH è ridotta, ci si potrebbe aspettare che l’espressione del PDK sia aumentata. In effetti questo è proprio ciò che è stato trovato. Premesso che l’enzima PDK esiste in 4 varianti negli esseri umani (PDK1, PDK2 etc), i norvegesi hanno trovato che PDK1, PDK2 e PDK4 sono sovraespressi nei pazienti ME/CFS (sia maschi che femmine), come indicato in figura 2. La maggiore differenza fra pazienti e controllo sano riguarda il PDK1 (p = 0.0018), quindi gli autori hanno indagato ulteriormente l’espressione di questo enzima, scoprendo che:
- il PDK1 è espresso maggiormente nelle donne che negli uomini (figura 3, M);
- il PDK1 è tanto più espresso quanto più severa è la patologia (figura 3, N);
- il PDK1 è espresso in modo maggiore in chi è malato da più di 10 anni (figura 3, O);
- il PDK1 è espresso maggiormente in chi compie meno di 2200 passi al giorno (figura 3, P).
In condizioni di salute, l’aumento della espressione genica dei PDK (con conseguente inibizione del PDH) si verifica durante periodi di mancanza di cibo. Quindi – in un certo senso – i pazienti ME/CFS soffrono le conseguenze della mancanza di cibo, pur vivendo nella disponibilità di risorse alimentari.

PPARD
Fluge e Mella hanno esaminato – sempre nelle PBMC – l’espressione di una serie di altri geni, tra cui i geni PPARD (Peroxisome proliferator-activated receptor) che svolgono moltissimi ruoli nella regolazione del metabolismo cellulare, tra cui la regolazione del metabolismo degli zuccheri, dei carboidrati e delle proteine (Dunning, KR et al. 2014). I PPAR sono tre, ma tra questi l’unica differenza significativa tra pazienti e controllo sano è nel PPARD, che è sovra espresso nei pazienti (vigura 4, F). E’ interessante notare che PPARD controlla l’espressione genica del PDK4: infatti agonisti del recettore PPARD aumentano l’espressione genica del PDK4, nelle cellule muscolari umane (Abbot EL et al. 2005). Questo significa che una possibile catena di eventi è questa:
- l’aumentata espressione di PPARD aumenta a sua volta l’espressione di PDK4, che inibisce il piruvato deidrogenasi.
Tuttavia questo è solo uno dei possibili percorsi regolatori coinvolti in questo complessi fenomeni.

Espressione di altri geni
Gli Autori norvegesi hanno indagato l’espressione di numerosi altri geni, trovando altre differenze fra controlli sani e pazienti. In particolare, il gene SIRT4 è sovra espresso (figura 5, K), così come il gene MPC1 e il gene per la subunità PDHA del PDH (figura 5, I e G). E’ interessante che questi tre geni siano strettamente legati fra loro, infatti la proteina codificata da SIRT4 inibisce la subunità PDHA del PDH, mentre MPC1 aumenta l’espressione genica di PDHA. Insomma, da un lato si inibisce questo pezzo del PDH e dall’altro si cerca di aumentarne il numero, per compensare!

Norvegia vs Pisa
Ho parlato più volte dell studio sulla espressione genica nei mitocondri dei pazienti ME/CFS della Reumatologia di Pisa (vedi qui). Ricordo che nella prima parte di quello studio si verificò l’espressione genica di 194 proteine mitocondriali in due gemelli, uno sano e uno con ME/CFS. In questo caso le cellule usate sono le piastrine, non le PBMC. Confrontando questi dati con quelli dello studio norvegese si osserva quanto segue.
- La subunità PDHA (PDH E1-alpha) è sovra espressa sia nello studio norvegese (figura 5, G) che nel gemello con ME/CFS, che ne esprime 1.6 volte di più del fratello sano (Tabella S1); il gemello malato sovraesprime anche la subunità E1 beta.
- la PDK1 e la PDK3 sono sovraespresse nel gemello malato (1.54 volte e 1.73 volte, rispettivamente) (Tabella S1). Nello studio norvegese ad essere sovraespresse sono la PDK1, la PDK2 e la PDK4 (figura 3), quindi esiste una discrepanza fra i due risultati.
Nel complesso si riscontrano dunque sia somiglianze che incongruenze fra questi due studi.
Cosa emerge?
Riassumo i punti salienti di quanto qui visto:
- i mitocondri consumano amminoacidi al posto dello zucchero per produrre energia, e questo fa pensare a un possibile blocco dell’enzima piruvato deidrogensi (PDH);
- una serie di enzimi che inibiscono il piruvato deidrogenasi (PDK1,2,4) sono effettivamente sovra espressi nei pazienti ME/CFS, rispetto ai controlli sani;
- una proteina (la PPARD) che aumenta l’espressione del PDK4 è sovra espressa nei pazienti ME/CFS;
- la proteina SIRT4, che inibische la subunità PDHA del PDH, è sovra espressa nei pazienti ME/CFS e, come forma di compenso, la subunità PDHA è sovra espressa, probabilemnte grazie alla sovra espressione di MPC1, che ha funzione regolatoria.
Detto in termini più semplici:
-
qualcosa induce l’espressione di una serie di meccanismi che riducono la funzione del piruvato deidrogenasi, costringendo i pazienti ME/CFS a bruciare amminoacidi al posto dello zucchero. Ma dei sistemi di compenso intervengono per cercare di riportare il metabolismo energetico alla normalità, senza riuscirci. I sistemi di compenso sono diversi fra maschi e femmine, ma il difetto metabolico a monte è lo stesso nei due sessi.
In un prossimo articolo riporterò la terza parte dello studio, che ha esaminato l’effetto del sangue dei pazienti su cellule umane in coltura. In quella sede torneremo sulle conclusioni da trarre da questi dati, e da quelli di altri studi.

6 thoughts on “Mitocondri norvegesi, secondo atto: espressione genica”